The Potential Role of Hemispheric Attentional Differentiation in a TBT Model and Voting Process
I find both the order of the timing of the acquisition of lateralization (predating mammals ) as well as the recognition that lateralization is a fundamental administrative difference of available information, very significant. (And also agree it does not make more primitive intelligence impossible, if we have no lateralization in place).
Also the fact, that the loss of a hemisphere early in life (when a single hemisphere can still learn to do the jobs of two hemisheres) is better in cognitive terms than losing a hemisphere later in life, when the functions were already split across both hemispheres, supports the notion that two fundamentally complimentary forms of attention are needed for what we consider to be a healthy cognitive balance.
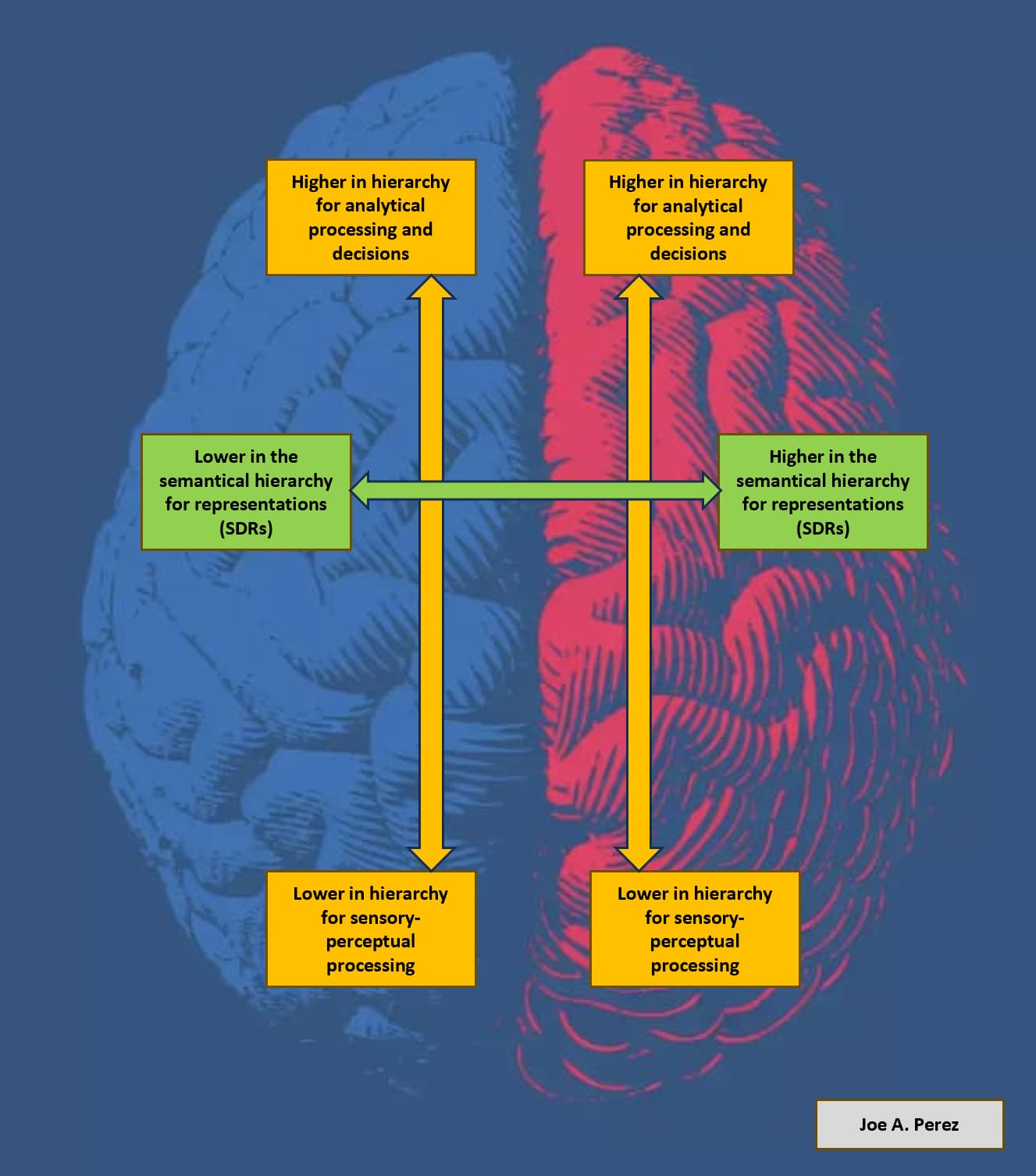
It must be noted, that hemispheric specialization is not essential for motor control which predates lateralization. Our left hemisphere on its own already masters the fine motoric control of a right hand in right-handed individuals and vice-versa in left-handed ones. So at an HTM-level of motor-sensory control, lateralization is not strictly needed, but for higher cognive “understanding” (disambiguation) of the object at the TBT-level of cognitive voting processes it may well be very fundamental. The left brain sees the elements of say an object or of a body of language (text) information. But it is the right brain that identifies the whole as a specific singular object, or in language it identifies the actual intention of the author of that language, rather than just seeing the literal meaning of that same text in the left-brain. It is only in the typical right brain that we see the singular identity of someone’s face, but in the left-brain we can consciously analyze the features of that face. The left is like Spock in Startrek and the right is like Captain Kirk. The left is reductionist and “conscious” of details. The right is holistic and conscious of specificity and of context. In my opinion this right side of our brains plays a fundamental role in the TBT voting process across the corpus collosum. Both hemispheres collaborate intensively, and as McGilchrist points out, the left hemisphere is barely aware of the right, but the right hemisphere is very aware of the left, because it integrates all the elements of output from the left. If we take this in conjunction with the functional differences of a PFC and the motor-sensory functions of our posterior brain, we can start to map out a more accurate layout of a TBT model.
The right brain may be more bottom-up cognitively and the left brain may be more top-down cognitively (in sequential terms of voting), but both have their control functions in the pre-frontal cortex and their sensory functions in the posterior sections of the brain.